|
Краткий курс анатомии и физиологии ЦНС
|
|
|
John_Smith
|
#16 | Среда, 02.07.2014, 11:36
|

Автор темы
Фильм
Юзер-бар +
Строение мозговой коры
Кора в основном состоит из 6 слоев:
1. Молекулярный слой – содержит небольшое количество мелких ассоциативных клеток;
2. Наружный зернистый слой – в его состав входят мелкие нейроны;
3. Пирамидный слой– самый широкий из всех слоев коры – состоит из малых и средних пирамидных
клеток
4. Внутренний зернистый слой – образован мелкими звездчатыми нейронами;
5. Ганглиозный слой– образован крупными пирамидными нейронами, наибольшего размера достигающих в
прецентральной извилине.
6. Слой полиморфных клеток – здесь находятся нейроны преимущественно веретенообразной формы. Этот
слой граничит с белым веществом.
Учение о структурных особенностях строения коры головного мозга носит название
архитектоники. Клетки коры больших полушарий менее специализированы, чем
нейроны отдельных отделов мозга, тем не менее, определенные их группы
анатомически и физиологически связаны с теми или иными специализированными
отделами мозга. Из 6 слоев клеток коры
верхние слои развиты у человека наиболее сильно. Нижние слои коры имеют связи с
периферическими рецепторами (IV слой) и с
мускулатурой (V слой) и носят название
«первичных» или «проекционных» корковых зон, вследствие их непосредственной
связи с периферическими отделами анализатора. Над «первичными» зонами
надстраиваются системы «вторичных» зон (II
и III слои), в которых преобладают
ассоциативные связи с другими отделами коры, поэтому они называются
проекционно-ассоциативными. Мозг как субстрат психических процессов представляет собой единую систему, единое
целое, состоящее, однако, из различных участков и зон, которые выполняют
различную роль в реализации психических функций. Морфологическая и функциональная неоднородности коры больших полушарий тесно связаны между собой.
По данным Института мозга, описаны следующие цито архитектонические поля коры
больших полушарий: постцентральная область (поля
1, 2, 3, 43), прецентральная (поля 4, 6),
лобная (поля 8-12, 44-47), теменная (поля 5, 7, 39, 40), височная (поля 20-22, 36-38, 41, 41), затылочная (17-19) и поясная область (поля 23-25, 31-33).Все данные (анатомические, физиологические и клинические) свидетельствуют о ведущей
роли коры больших полушарий в мозговой организации психических процессов.В нейропсихологии на основе анализа нейропсихологических данных (т.е. изучения нарушений психических процессов при различных локальных поражениях
мозга) была разработана общая структурно-функциональная модель мозга как
субстрата психической деятельности. Эта
модель,предложенная А. Лурия, характеризует наиболее общие закономерности работы мозга как единого целого и является основой для
объяснения его интегративной деятельности. Согласно данной модели весь мозг
может быть подразделен на три основных структурно-функциональных блока: а)
энергетическии блок, или блок регуляции уровней активности мозга; б) блок
приема, переработки и хранения экстероцептивной (исходящей извне) информации;
в) блок программирования, регуляции и контроля над протеканием психической деятельности.
Каждая высшая психическая функция осуществляется при участии всех трех блоков,
каждый из которых вносит свой вклад в ее реализацию. Блоки характеризуются
определенными особенностями строения, физиологическими принципами, лежащими в
основе их работы, и той ролью, которую они играют в осуществлении психических
функций.Первый энергетический блок регулирует два типа процессовактивации: общие генерализованные изменения активации мозга, являющиеся основой
различных функциональных состояний, и локальные избирательные активационные
изменения, необходимые для осуществления высших психических функций. Функциональное значение первого блока в обеспечении психических функций, прежде всего,
состоит, как уже говорилось выше, в регуляции процессов активации, â обеспечении того общего активационного
уровня, на котором разыгрываются все психические функции, в поддержании общего
тонуса ЦНС, необходимого любой психической деятельности. Этот аспект работы
первого блока имеет непосредственное отношение к процессам внимания, а также
сознания в целом. Помимо общих неспецифических активационных функций первый
блок мозга непосредственно связан с процессами памяти, с запечатлением,
хранением и переработкой разно модальной информации. Второй блок - блок приема, переработки è хранения
информации включает в себя основные анализаторные системы: зрительную, слуховую
и кожно-кинестетическую, корковые отделы (зоны) которых расположены в задних
отделах больших полушарий. Все три анализаторные системы организованы по общему принципу: они состоят из
периферического (рецепторного), проводникового и центральных отделов. Проводниковый
отдел анализаторов обычно включает несколько уровней. Центральный отдел
анализаторов - кора больших полушарий. В целом анализаторы - это аппараты, подготавливающие ответы организма на внешние
раздражители. Кора задних отделов больших полушарий обладает рядом общих черт, позволяющих
объединить различные уровни анализаторных систем в единый блок мозга. Â коре задних отделов мозга выделяют первичные,
вторичные и третичные поля, а по терминологии И.П. Павлова - «ядерные зоны
анализаторов» и «периферию». В ядерную зону зрительного анализатора входят 17, 18 и 19-е поля, в ядерную зону
кожно-кинестетического анализатора - 3, 1,
2-е, частично 5-е поле, в ядерную зону звукового анализатора - 41, 42 и 22-е поля, из них первичными полями
являются - 17, 3 и 41-е. Остальные - вторичные.
Вторичные корковые поля осуществляют синтез раздражителей, функциональное объединение
различных анализаторных зон, принимая непосредственное участие в обеспечении
различных гностических видов психической деятельности. Третичные поля коры больших полушарий находятся вне «ядерных зон» анализаторов. К ним относятся верхнетеменная область (7 и 40), нижнетеменная (39), средневисочная область (21
и частично 37). Функциональное значение
третичных полей коры многообразно. С их участием осуществляются сложные
надмодальностные виды психической деятельности - символической, речевой,
интеллектуальной.Третийструктурно-функциональный блок мозга - блок
программирования, регулирования и контроля над протеканием психической
деятельности - включает моторные,
премоторные и префронтальные отделы коры лобных отделов головного мозга. Кора
лобных долей мозга занимает 24% поверхности
больших полушарий. В лобной коре выделяют моторную кору (4, 6) и немоторную (9, 10, 11, 12,
46, 47). Эти области имеют различное строение и функции. Моторная
лобная кора составляет ядерную зону двигательного анализатора .Общая структурно-функциональная модель организации мозга, предложенная А.Р. Лурия,
предполагает, что различные этапы произвольной, опосредованной, осознанной
психической деятельности осуществляются с обязательным участием всех трех
блоков мозга.Согласно современным психологическим представлениям, каждая психическая деятельность имеет строго определенную структуру: она начинается с фазы
мотивов, намерений, замыслов, которые затем превращаются в определенную
программу деятельности, включающую «образ результата» и представления о
способах реализации этой программы, а затем реализуется с помощью определенных
операций. Завершается психическая деятельность этапом сличения полученных результатов ñ исходным «образом результата». В случае несоответствия
полученных данных психическая деятельность продолжается до получения нужного
результата. Поражение любого из трех блоков (или нарушение каких-либо отделов
этих блоков) отражается на любой психической деятельности, так как приводит к
нарушению соответствующей стадии или этапа ее реализации. Данная общая схема
формирования мозга как субстрата сложных сознательных форм психической
деятельности находит конкретное подтверждение при анализе различных нарушений
высших психических функций, возникающих вследствие локальных поражений
головного мозга.
Почему вы так волнуетесь? У вас прекрасная болезнь, болезнь Пушкина.
|
Статус: нет меня
|


|
|
|
John_Smith
|
#17 | Среда, 02.07.2014, 11:39
|
Автор темы
Фильм
Юзер-бар +
Белое вещество полушарий
К белому веществу полушария относятся внутренняя капсула, свод и волокна,
проходящие через спайки мозга, а также
волокна, соединяющие в полушарии отдельные участки коры между собой, кору и
базальные ядра. Нервные волокна в полушариях делят на три группы:ассоциативные, комиссуральные и проекционные.
Ассоциативные волокна связывают между собой участки одного и того же полушария. Короткие
волокна соединяют соседние извилины, а длинные волокна – доли полушария. Комиссуральные волокна связывают симметрично расположенные участки разных полушарий, образуя
при этом спайки. К ним относятся: мозолистое тело, передняя спайка мозга и
спайка свода. Проекционные волокна связывают кору полушарий мозга с нижележащими отделами головного и
спинного мозга. При этом выделяют восходящие и нисходящие волокна. Проекционные
волокна, находящиеся ближе всего к коре, образуют лучистый венец. Все они затем
сходятся во внутреннюю капсулу, представляющую собой слой белого вещества,
расположенный между хвостатым ядром, чечевицеобразным ядром и зрительным
бугром. На горизонтальном разрезе капсула имеет вид бумеранга, открытого в
латеральную сторону. Свод находится под мозолистым телом.В области средней трети мозолистого тела свод, раздваиваясь, дугообразно
загибается впереди вниз. При этом образуются столбы свода, которые
заканчиваются в мамиллярных телах гипоталамуса. Кроме того, раздваиваясь кзади,
свод образует ножки свода, которые идут в нижний рог бокового желудочка. Между
столбами свода и ножками свода натянута тонкая прозрачная перегородка.
Почему вы так волнуетесь? У вас прекрасная болезнь, болезнь Пушкина.
|
Статус: нет меня
|
|
|
|
John_Smith
|
#18 | Среда, 02.07.2014, 11:42
|
Автор темы
Фильм
Юзер-бар +
Проводящие пути
Проводящиепути – это пучки нервных волокон,проходящих в определенных зонах белого вещества головного и спинного мозга,
объединенных общностью морфологического строения и функции, и соединяющих между
собой нервные центры. В ЦНС выделяют восходящие и нисходящие проводящие пути.
Ввиду их большого количества мы рассмотрим только основные.
Восходящие пути.
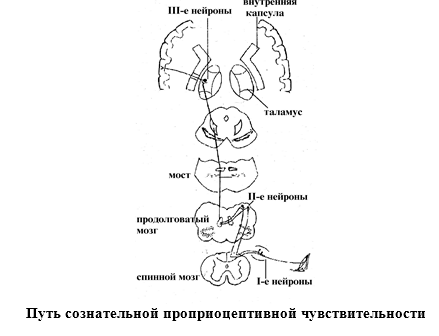
Путь сознательной проприоцептивной чувствительности (путь Голля и Бурдаха).Первые нейроны этого пути лежат в спинальном ганглии. Причем от 19 нижних
ганглиев идут волокна, образующие пучок Голля, а от 12 верхних – пучок
Бурдаха. Аксоны первых нейронов проходят
в составе задних корешков и, не заходя в серое вещество задних рогов, проходят
в задние канатики, поднимаются вверх и оканчиваются в продолговатом мозге в
нежном ядре и клиновидном ядре, соответственно. В этих ядрах лежат тела вторых нейронов. Их аксоны совершают перекрест и в
составе медиальной петли проходят транзитом через мост, средний мозг и
заканчиваются в латеральных ядрах таламуса. Здесь располагаются тела третьих
нейронов. Их аксоны проходят через внутреннюю капсулу и оканчиваются в задней
центральной извилине.
Почему вы так волнуетесь? У вас прекрасная болезнь, болезнь Пушкина.
|
Статус: нет меня
|
|
|
|
John_Smith
|
#19 | Среда, 02.07.2014, 11:43
|
Автор темы
Фильм
Юзер-бар +
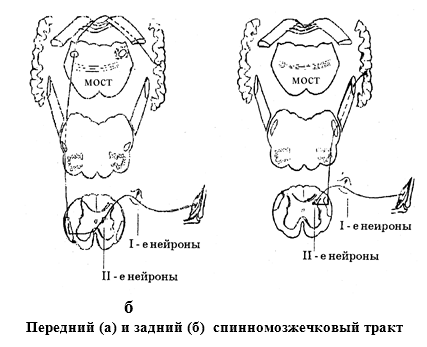
Пути бессознательной проприоцептивной чувствительности.1) Передний спинномозжечковый путь (путь Говерса). Первые нейроны этого пути находятся в спинальном ганглии. Их аксоны
входят в составе задних корешков в боковые рога серого вещества спинного мозга.
Здесь лежат тела вторых нейронов. Их аксоны переходят на другую половину
спинного мозга в боковые канатики (первый перекрест). Они проходят транзитом
через продолговатый мозг, ретикулярную формацию моста и, достигнув верхнего
мозгового паруса, совершают второй перекрест. После этого волокна по верхним
ножкам мозжечка достигают коры червя, где и заканчиваются.
2)Задний спинно мозжечковый путь. Первые нейроны пути находятся в спинальном
ганглии. Их аксоны входят в составе задних корешков в задние рога серого вещества
спинного мозга. Здесь лежат тела вторых нейронов. Их аксоны входят в состав
боковых канатиков своей стороны и поднимаются вверх до продолговатого мозга и
по нижним мозжечковым ножкам достигают коры червя.
Почему вы так волнуетесь? У вас прекрасная болезнь, болезнь Пушкина.
|
Статус: нет меня
|
|
|
|
John_Smith
|
#20 | Среда, 02.07.2014, 11:45
|
Автор темы
Фильм
Юзер-бар +
Экстероцептивные проводящие пути.
1) Боковой спиноталамический путь – обеспечивает болевую и температурную чувствительность. Первые нейроны пути находятся в спинальном ганглии. Их аксоны
входят в составе задних корешков в задние рога спинного мозга. Здесь лежат тела
вторых нейронов. Их аксоны поднимаются 1) выше на 2-3 сегмента и, совершая перекрест, выходят в боковой канатик
противоположной стороны, далее идут транзитом через продолговатый мозг, мост и
средний мозг в составе медиальной петли и оканчиваются в латеральных ядрах
таламуса. Здесь находятся тела третьих нейронов. Их аксоны проходят через
внутреннюю капсулу и оканчиваются в задней центральной извилине.
2) Передний спиноталамический путь – обеспечивает ощущения осязания и давления. Первые
нейроны пути находятся в спинальном ганглии. Их аксоны входят в составе задних
корешков в задние рога спинного мозга. Здесь лежат тела вторых нейронов. Их
аксоны, совершая перекрест, входят в передний канатик противоположной стороны и
в его составе направляются вверх, идут транзитом в составе медиальной петли,
через продолговатый мозг и заканчиваются в латеральных ядрах таламуса. Здесь
находятся тела третьих нейронов. Их аксоны проходят через внутреннюю капсулу и
оканчиваются в задней центральной извилине. 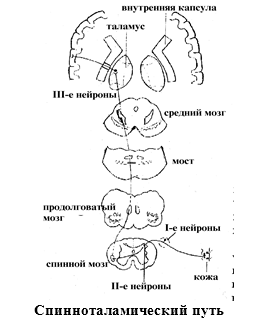
Почему вы так волнуетесь? У вас прекрасная болезнь, болезнь Пушкина.
|
Статус: нет меня
|
|
|
|
John_Smith
|
#21 | Среда, 02.07.2014, 11:47
|
Автор темы
Фильм
Юзер-бар +
Нисходящиепути.
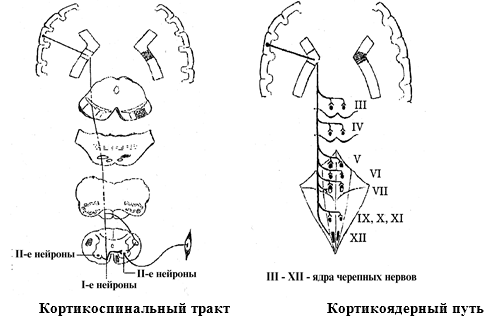
Пирамидные пути. 1) Кортикоспинальный тракт. Тела первых нейронов – гигантские пирамидные клетки
Беца, располагающиеся в верхних двух третях передней центральной извилине. Их
аксоны проходят через внутреннюю капсулу
и идут транзитом через ножки мозга, мост, продолговатый мозг. В продолговатом
мозге волокна образуют выступающие вперед валики, пирамиды.
На границе со спинным мозгом часть волокон совершает перекрест и в виде бокового кортиокспинального тракта спускается в боковые
канатики спинного мозга. Эти волокна заканчиваются на своей стороне в
двигательных ядрах передних рогов спинного мозга. Не перекрещенная часть волокон,
образуя передний кортикоспинальный тракт, идет в передних канатиках
шейно-грудного отдела спинного мозга, совершает посегментный перекрест и
заканчивается в двигательных ядрах передних рогов противоположной стороны. Здесь располагаются тела вторых нейронов. Аксоны переднего пирамидного пути проводят
импульсы к мышцам туловища, а бокового – к мышцам конечностей
.
2) Кортико ядерный путь. Первые нейроны – гигантские пирамидные клетки Беца нижней части передней центральной извилины. Их аксоны проходят через внутреннюю капсулу и идут
транзитом через ножки мозга, мост, продолговатый мозг. Здесь аксоны
заканчиваются на соматических двигательных ядрах черепномозговых нервов своей и
противоположной стороны (кроме ядра подъязычного нерва и нижней части ядра лицевого
нерва). В этих ядрах располагаются тела вторых нейронов. Их аксоны в составе
периферических черепных нервов идут к мышцам шеи, лица и головы
Почему вы так волнуетесь? У вас прекрасная болезнь, болезнь Пушкина.
|
Статус: нет меня
|
|
|
|
John_Smith
|
#22 | Среда, 02.07.2014, 11:51
|
Автор темы
Фильм
Юзер-бар +
Экстрапирамидные пути
1)Красноядерноспинальный тракт. Первые нейроны располагаются в красном ядре
среднего мозга. Их аксоны при выходе из покрышки среднего мозга совершают
перекрест, после которого транзитом проходят через ретикулярную формацию моста,
продолговатого мозга, спускаются в боковые канатики спинного мозга, доходят до
своего сегмента и заканчиваются в двигательных ядрах передних рогов. Здесь находятся тела вторых нейронов. Их аксоны в составе передних корешков выходят
из спинного мозга и далее в составе спинномозговых нервов достигают
поперечнополосатой мускулатуры.
2)Оливоспинальный тракт. Первые нейроны располагаются в нижней оливе
продолговатого мозга. Их аксоны, не перекрещиваясь, идут в составе передних
канатиков спинного мозга до двигательных нейронов передних рогов. Здесь
располагаются тела вторых нейронов, чьи аксоны достигают скелетных мышц.
3)Вестибулоспинальный и ретикулоспинальный тракты. Первые нейроны располагаются в
вестибулярных ядрах и ядрах ретикулярной формации, соответственно. Их аксоны,
не перекрещиваясь, идут в составе передних канатиков спинного мозга до
двигательных нейронов передних рогов. Здесь располагаются тела вторых нейронов,
чьи аксоны достигают скелетных мышц.
4)Тектоспинальный тракт. Первые нейроны располагаются в четверохолмии среднего
мозга. Их аксоны совершают перекрест, транзитом идут в передние канатики
спинного мозга, и заканчиваются на двигательных нейронах передних рогов. Здесь
располагаются тела вторых нейронов, чьи аксоны достигают скелетных мышц.
Функции нисходящих путей. Пирамидные и экстрапирамидные пути совместно обеспечивают регуляцию целенаправленных двигательных реакций.
При этом функции пирамидной системы модулируются деятельностью экстрапирамидной
системы. Схема взаимодействия этих функциональных систем ЦНС выглядит следующим образом. При
выполнении целенаправленного движения
общее направление движения обеспечивается регуляторными механизмами
экстрапирамидной системы. Она определяет пластический тонус мышц и обеспечивает
сокращение мышечных групп, участвующих в движении. Завершение целенаправленного
движения (например, захват предмета пальцами) осуществляется под контролем
пирамидной системы и ее нисходящих путей. Нарушение взаимодействия пирамидной и
экстрапирамидной систем при патологии ЦНС приводит к поражению двигательных
реакций организма. Экстрапирамидные пути осуществляют сложные безусловно рефлекторные реакции организма на
раздражение. Например, тектоспинальный тракт обеспечивает ориентировочный
рефлекс на неожиданные зрительные и слуховые раздражители; ретикулоспинальный
тракт тормозит двигательные реакции спинного мозга.
Почему вы так волнуетесь? У вас прекрасная болезнь, болезнь Пушкина.
|
Статус: нет меня
|
|
|
|
John_Smith
|
#23 | Среда, 02.07.2014, 11:53
|
Автор темы
Фильм
Юзер-бар +
Боковые желудочки мозга
Боковые желудочки являются полостью конечного мозга. Они залегают симметрично по обеим
сторонам от средней линии ниже уровня мозолистого тела. В каждом желудочке выделяют центральную часть, передний, задний и нижние рога. Центральная часть соответствует теменной доле полушария и представляет собой горизонтально
расположенное щелевидное пространство. Передний рог бокового желудочка располагается в лобной доле, задний – в затылочной, нижний
– в височной доле.
В центральную часть и нижний рог желудочка вдается сосудистое сплетение
Почему вы так волнуетесь? У вас прекрасная болезнь, болезнь Пушкина.
|
Статус: нет меня
|
|
|
|
John_Smith
|
#24 | Среда, 02.07.2014, 11:55
|
Автор темы
Фильм
Юзер-бар +
Сообщения желудочков головного мозга
1. Боковые желудочки через межжелудочковые отверстия сообщаются с IIIжелудочком.
2. III желудочек,кроме вышеописанных боковых, сообщается через водопровод мозга с IV желудочком.
3. IV желудочек,кроме третьего, сообщается благодаря боковым и медиальному отверстиям с
подпаутинным пространством головного мозга и с центральным каналом спинного
мозга. Заращение этих отверстий ведет к накоплению жидкости в подпаутинном пространстве
– водянке головного мозга.
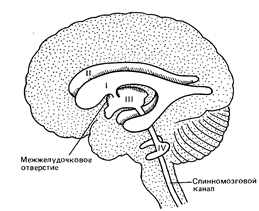
Желудочки головного мозга. Боковые желудочки (первыйи второй) расположены в самой середине каждого полушария; третий желудочек
занимает центральную часть большого мозга. Он сообщается с боковыми желудочками
(через межжелудочковые, или монроевы, отверстия) и с четвертым желудочком, находящимся
в стволе.
Почему вы так волнуетесь? У вас прекрасная болезнь, болезнь Пушкина.
|
Статус: нет меня
|
|
|
|
John_Smith
|
#25 | Среда, 02.07.2014, 11:56
|
Автор темы
Фильм
Юзер-бар +
Оболочки головного мозга
Головной мозг, как и спинной, окружен тремя соединительнотканными оболочками: наружной
твердой, средней паутинной и внутренней сосудистой.
Твердая оболочка. Ее наружная поверхность прилежит непосредственно к костям черепа, для которых является внутренней
надкостницей. С костями свода черепа твердая оболочка связана слабо и легко
отделяется, а с костями основания черепа – прочно. По определенным линиям твердая оболочка расщепляется на два листка, образуя синусы
(пазухи), выстланные эндотелием. В синусы впадают вены, по которым происходит
отток крови из мозга. Основной отток крови из синусов идет через внутренние
яремные вены. Синусы отличаются от обычных вен тем, что из-за наличия жестких
стенок не спадаются и не изменяют свой просвет при изменениях внутричерепного
давления. Со своей внутренней стороны твердая оболочка образует отростки, проходящие в виде
пластинок в щели между отдельными частями головного мозга и отделяющие эти
части друг от друга.
Паутинная оболочка головного мозга имеет вид тонкой прозрачной, но плотной пластинки, бедной сосудами и нервами. От твердой
оболочки она отделяется субдуральным пространством, а от мягкой – подпаутинным
пространством. Подпаутинное пространство заполнено спинномозговой жидкостью.
Паутинная оболочка соединяется с мягкой оболочкой с помощью соединительно тканных
перекладин.
Подпаутинное пространство не представляет общей полости одинаковой глубины, а состоит из
множества сообщающихся между собой щелей. Там где подпаутинная оболочка
перекидывается через глубокие борозды, образуются так называемые подпаутинные
цистерны. Все цистерны сообщаются друг с другом, а в области большого затылочного отверстия
подпаутинное пространство головного мозга сообщается с подпаутинным
пространством спинного мозга. От наружной поверхности паутинной оболочки отходят специфические выросты,
вдающиеся в венозные синусы твердой оболочки головного мозга. Это так называемые
паутинные грануляции, через которые происходит отток спинномозговой жидкости из подпаутинного пространства в
венозные синусы.
Мягкая (сосудистая) оболочка головного мозга плотно прилежитк веществу мозга. Она богата нервными и кровеносными сосудами и имеет большое
значение в питании мозга. В определенных местах мягкая оболочка проникает в
желудочки мозга и формирует складки, содержащие большое количество кровеносных
сосудов. Это сосудистые сплетения, продуцирующие из крови спинномозговую
жидкость.
Почему вы так волнуетесь? У вас прекрасная болезнь, болезнь Пушкина.
|
Статус: нет меня
|
|
|
|
John_Smith
|
#26 | Среда, 02.07.2014, 11:59
|
Автор темы
Фильм
Юзер-бар +
Вегетативная нервная система
Вегетативнуюнервную систему подразделяют на
симпатическую и парасимпатическую. Работа этих двух систем-антагонистов
поддерживает в организме стабильность внутренней среды
перед лицом вечно изменяющегося
мира. Главное различие между симпатической и парасимпатической системами заключается в том,
что первая мобилизует организм для
действия (катаболизм), а другая -
восстанавливает запасы энергии в организме (анаболизм). Основная функция симпатической системы - это мобилизация всего организма при чрезвычайных, экстремальных обстоятельствах. Такая мобилизация связана с рядом
сложных реакций, начиная с расщепления гликогена в печени (образующаяся при
этом глюкоза служит добавочным источником энергии) и кончая изменениями в циркуляции крови. Каждую из этих реакций, осуществляемых симпатической нервной
системой, легко понять как механизм приспособления к «аварийным» ситуациям,
выработанным в ходе эволюции. Обеспечение доступа к запасам энергии дает
организму максимум физических возможностей в непредвиденных ситуациях.
Уменьшение кровотока около поверхности тела снижает вероятность обильного
кровотечения при повреждении кожи,
тогда как усиленная подача крови
к глубже лежащим мышцам позволяет
развить большее физическое усилие. Кеннон назвал весь этот комплекс изменений
«реакцией борьбы и бегства». Его теоретические соображения о роли этой реакции
явились существенным стимулом для развития психофизиологии и современных
представлений об «общей активации» организма. Следующее по важности различие - то, что симпатическая система имеет тенденцию
действовать быстро и как единое целое, тогда как парасимпатическая активация
более кратковременна и носит локальный характер. Действие симпатической системы
обычно проявляется диффузно (охватывает весь организм) и поддерживается относительно
долго. С другой стороны, действие парасимпатической системы, способствующее
сохранению и поддержанию основных ресурсов организма, локально и относительно
кратковременно.Эффектысимпатической и парасимпатической систем на органы и системы организма
противоположны друг другу. В то время как симпатическая нервная система
ускоряет сокращения сердца, парасимпатическая их замедляет, она усиливает также
приток крови к желудочно-кишечному тракту и стимулирует превращение глюкозы в
гликоген печени. Большинство, но не все внутренние органы получают иннервацию
от обеих систем. Поскольку обе они работают согласованно, трудно бывает определить, связано ли данное
изменение функции с активностью той или другой из них. Например, замедление
ритма сердца может указывать и на усиленную активность парасимпатической
системы, и на ослабление действия ее антагониста.
Последнее различие в функционировании обеих систем связано с особенностями их структурной
организации. В соматической нервной системе каждый нейрон, тело которого
находится в ЦНС, имеет длинный отросток – аксон, проводящий нервные импульсы к
органу-мишени. В произвольной мускулатуре такой аксон образует синапс в области
двигательной пластинки мышечного волокна. Соматическая нервная система, таким
образом, имеет «однонейронный путь». В вегетативной же системе путь к
органу-исполнителю двухнейронный. Место соединения между этими двумя нейронами находится
в вегетативном ганглии. Симпатические волокна выходят из средней части спинного мозга - из грудного и поясничного отделов, поэтому симпатическую систему иногда называют тораколюмбальной. Ее аксоны сходятся к группе симпатических
ганглиев, расположенных с обеих сторон спинного мозга. В этих ганглиях с плотно
расположенными нейронами существуют большие
возможности для электрических «переключений», и импульс,
пришедший из любого участка симпатической системы, может вызвать активацию всей
этой системы. Парасимпатические волокна образуют синапсы недалеко от
иннервируемого органа, они выходят из спинного мозга выше или ниже места выхода
симпатических волокон - из черепного и кресцового отделов. В связи с этим парасимпатическую
нервную систему еще называют краниосакральной. Ее ганглии расположены далеко
друг от друга, и поэтому нервные импульсы
оказываются более специфическими. Кроме этого симпатическая нервная
система иннервирует надпочечники, которые при стимуляции выделяют адреналин и норадреналин.
Норадреналин гормонального происхождения попадает в симпатические синапсы и
усиливает их действие. Для распада этого гормона требуется некоторое время, поэтому для прекращения
активности симпатической системы также требуется некоторое время. Для парасимпатической
системы существует другой медиатор - ацетилхолин. В синапсах этой системы
ацетилхолин быстро инактивируется холинэстеразой, в связи с этим
парасимпатические эффекты четко ограничены не только в пространстве, но и во
времени. Важным исключением из этого правила являются симпатические волокна,
иннервирующие потовые железы - они активируются ацетилхолином. Таким образом,
различия между катаболическим действием симпатической системы и анаболическим
действием парасимпатической системы обусловлены особенностями их анатомического
строения. Нервные механизмы регуляции вегетативных функций имеют иерархическую структуру. Первым
уровнем этой иерархии являются внутри органные периферические рефлексы,
замыкающиеся в интрамуральных ганглиях вегетативной нервной системы, средним
уровнем являются нервные центры ствола
мозга, высшим уровнем - кора больших полушарий. Она обеспечивает координацию
вегетативных и соматических функций в сложных поведенческих реакциях организма,
возникновение которых обусловлено индивидуальным опытом.
Установлено, что в продолговатоммозге расположены центры, тормозящие деятельность сердца, возбуждающие слезоотделение и секрецию слюнных и желудочных желез, поджелудочной железы,
вызывающие выделение желчи из желчного пузыря и желчного протока, возбуждающие
сокращение
желудка и тонкого кишечника. Здесь же, в ретикулярной формации находится
сосудодвигательный центр, координирующий и интегрирующий деятельность нейронов
симпатического отдела нервной системы, расположенных в тораколюмбальных сегментах
спинного мозга и посылающих на периферию сосудосуживающие импульсы. Характерной особенностью сосудодвигательного центра продолговатого мозга и нейронов
блуждающего нерва, тормозящих сердечную деятельность, является то, что они находятся постоянно в
состоянии автоматической активности (тонуса), в результате чего артерии и
артериолы всегда несколько сужены, а сердечная деятельность замедлена.Приучастии нейронов ядер блуждающих нервов осуществляется различные рефлексы на
сердце, в том числе рефлекс Гольца, глазосердечный (рефлекс Ашнера), дыхательно-сосудистый
и др .Многие рефлекторные реакции сердца осуществляются сопряженно с изменениями сосудистого
тонуса. Это обусловлено связями,
существующими между нейронами, регулирующими деятельность сердца и сосудистый
тонус. Импульсы к спинномозговым нейронам симпатической системы, иннервирующим сосуды, передаются от
сосудовигательного центра по ретикулоспинальным путям. Сосудорасширяющие
рефлексы сосудодвигательного центра имеют, как правило, регионарный характер,
сосудосуживающие - охватывают обширные области тела. В отличие от дыхательного центра, центры регуляции сердечной деятельности и
сосудистого тонуса, хотя и находятся под влиянием коры полушарий мозга, обычно
не могут быть произвольно возбуждены или заторможены (для этого требуется
специальная тренировка). Рефлекторные центры продолговатого мозга, регулирующие деятельность пищеварительных органов,
осуществляют свое влияние через парасимпатические нервные волокна,
приходящие к слюнным железам в составе
языкоглоточного и лицевого нервов, а к желудку, поджелудочной железе, тонкому
кишечнику, желчному пузырю и желчным протокам - в составе блуждающего нерва.
В среднем мозге, в передних буграх четверохолмия, находятся нервные центры зрачкового
рефлекса и аккомодации глаза. Деятельность вегетативных центров, расположенных в спинном, продолговатом и среднем мозге, в
свою очередь регулируется высшими вегетативными центрами гипоталамуса.
Особенность ответных реакций, возникающих при раздражении разных участков гипоталамуса,
заключается в том, что в них участвуют многие органы тела. Эти реакции являются
комплексными и интегрированными. Ядра гипоталамуса принимают участие во многих
общих, в том числе поведенческих, реакциях. Так, гипоталамус участвует в половых
и агрессивно-оборонительных реакциях. Точечное раздражение его
вентромедиального ядра вызывает у кошки резко выраженный агрессивный эффект -
так называемую реакцию мнимой ярости.Деятельность гипоталамуса в свою очередь контролируется высшими отделами ЦНС - подкорковыми ядрами, мозжечком и корой больших полушарий, с которыми
гипоталамус связан как прямыми нервными путями, так и путями, проходящими в
составе ретикулярной формации мозгового ствола
Оказывая активирующее и тормозящее влияние на
различные отделы ЦНС, ретикулярная формация повышает активность вегетативных нервных центров. Она оказывает на них тонизирующее влияние. Поэтому симпатический отдел может рассматриваться в
функциональном единстве с ретикулярной формацией.Экспериментальнопоказано, что введение адреналина повышает тонус ретикулярной формации, в
результате чего усиливается ее активирующее влияние на большие полушария .На вегетативную нервную систему существенное влияние оказывает мозжечок. При его удалении возникает угнетение моторной, в частности периодической, деятельности пищеварительного
тракта и секреторной функции желез желудка и кишечника.
Как известно, подкорковые ядра, в частности полосатое тело, участвуют в осуществлении сложных,
безусловно-рефлекторных реакций организма, которые включают и вегетативные
компоненты. Вегетативные реакции могут формироваться при возбуждении
подкорковых ядер вследствие того, что последние имеют прямые связи с
ретикулярной формацией мозгового ствола и гипоталамусом. Важную роль в регуляции деятельности внутренних органов имеют нервные образования,
которые входят в состав лимбической системы, или висцерального мозга:
гиппокамп, поясная извилина, миндалевидные ядра.
Лимбическая система участвует в формировании эмоций и таких поведенческих реакций, в осуществлении которых имеет место ярко
выраженный вегетативный компонент. Влияние висцерального мозга на функции
органов, иннервируемых вегетативной
нервной системой, осуществляется через гипоталамус.
Высшим уровнем регуляции вегетативной нервной системы является кора полушарий мозга. Впервые В.Я. Данилевским в 1874 г. было установлено, что раздражение некоторых участков коры больших полушарий у собак
вызывает изменения дыхания, сердечной деятельности, сосудистого тонуса.
Впоследствии В.М. Бехтерев, Н.А. Миславский, другие физиологи и нейрохирурги
наблюдали изменения многих вегетативных функций при раздражении разных участков
коры больших полушарий. Школа И.П. Павлова рассматривает нейроны коры больших полушарий, участвующих в
регуляции функций внутренних органов, как корковое представительство интероцептивного
анализатора. В регуляции вегетативных функций большое значение имеют лобные
доли коры больших полушарий. Раздражение этих долей вызывает изменение дыхания,
пищеварения, кровообращения и половой деятельности. Поэтому считается, что в
передних отделах коры больших полушарий находятся высшие центры вегетативной
нервной системы.В коре больших полушарий существуют зоны, связанные нисходящими путями с ретикулярной формацией ствола мозга. Эти зоны расположены в сенсомоторной коре,
лобных глазодвигательных полях, поясной извилине, верхней височной извилине и
околозатылочной области. По нисходящим путям, идущим от этих зон коры, импульсы
поступают к ретикулярной формации, а от нее - к гипоталамусу и гипофизу. Имеются
также прямые пути, идущие от лобной доли и от поясной извилины
к гипоталамусу. С помощью этих путей кора оказывает регулирующее влияние на
нижележащие уровни вегетативной нервной системы .Вегетативная нервная система регулирует работу сердца, желез и непроизвольной (гладкой)
мускулатуры без активного участия нашего сознания. В течение многих лет
считалось, что функции вегетативной системы недоступны для нормального
самоконтроля. Недавние эксперименты с созданием обратной связи и изучение
практики восточных мистиков с их древней религиозной традицией тренировки тела
позволяют предполагать, что и так называемую «непроизвольную» мускулатуру можно
поставить под контроль воли. Влияние коры головного мозга на многие внутренние органы доказано в опытах с воздействием
на человека гипнотического внушения. Например, показано, что внушением можно
вызвать учащение или замедление деятельности сердца, расширение или сужение
сосудов, усиление отделения мочи почками, выделение пота, изменение
интенсивности обмена веществ. Известны случаи, когда влияние коры полушарий мозга проявлялось настолько резко, что
человек мог произвольно вызывать увеличение частоты сердечных сокращений,
поднятие волос и появление «гусиной» кожи, обычно наблюдаемой в результате
охлаждения тела, а также изменять ширину зрачков, зависящую от тонуса гладких
мышц радужки глаза. Однако,если бы протекание внутренних функций требовало нашего повседневного контроля,
у нас почти не оставалось бы времени на что-либо иное, кроме непрерывных забот
о поддержании жизнедеятельности собственного организма. Специализация и
автоматический режим работы вегетативной нервной системы создали возможность
для дальнейшей эволюции мозговых механизмов, обеспечивающих высшие психические
функции.
Почему вы так волнуетесь? У вас прекрасная болезнь, болезнь Пушкина.
|
Статус: нет меня
|
|
|
|
John_Smith
|
#27 | Среда, 02.07.2014, 12:06
|
Автор темы
Фильм
Юзер-бар +
Электрическая активность мозга (электроэнцефалография)
Мозг состоит из более чем 10 млрд клеток и каждая из них представляет собой миниатюрную станцию, способную в
возбужденном состоянии создавать электрический потенциал. Впервые эта
электрическая активность была зарегистрирована в 1875 г. английским хирургом
Ричардом Ксйтоном. Он впервые показал, что у животного можно зарегистрировать
электрическую активность мозга. В своих экспериментах этот исследователь
регистрировал фоновую активность с поверхности сенсорной коры мозга кролика. Приблизительно через 50 лет (1924) сходные наблюдения были сделаны на человеке австрийским
психиатром Хансом Бергером. Запись мозговых волн осуществлялась с помощью
специального прибора, способного регистрировать и усиливать потенциалы,
создаваемые нервными клетками. Это делалось с помощью электродов, прикрепленных
к коже черепа испытуемого. Регистрируемые слабые потенциалы усиливались и
отображались графически в виде волн, которые Бергер назвал "мозговые
волны". В ходе своих исследований он установил, что часть этих потенциалов
принадлежит мозгу, а не обусловлена активностью мышц головы; что электрические
характеристики этих сигнапов зависят от состояния испытуемых; он выделилосновные виды электрической активности мозга: апьфа-волны и бета-волны; показал
что электрическая активность мозга может рассматриваться в качестве индикатора
общей активности мозга. Метод регистрации электрической активности мозга
получил название электроэнцефалографии (ЭЭГ).Последующие исследования показали,что показатели ЭЭГ качественно отличаются от открытых ранее более простых
показателей активности вегетативной нервной системы. Периодические волны
электрической активности, возникающие при сокращении сердца, - это сама
простота по сравнению с ужасающей сложностью ЭЭГ. Сложность формы волн ЭЭГ как
будто бросает вызов исследователям, пытающимся найти в них хоть какой-нибудь
смысл.Современники Бергера отнеслись кего сообщениям скептически и "мозговые воины" стали общепризнанным
фактом только после того, как Эндриан и Мэттьюз осуществили наглядную
демонстрацию записи ЭЭГ на заседании английского физиологического общества в
1935 г .Последующие годы были волнующими для исследователей, которые с энтузиазмом проникали в тайны глубин мозга.
Многочисленные работы позволили установить электро энцефалографические корреляты эпилепсии:записи с разных участков черепа регестрируют колебания разной формы:- при опухолях мозга в окружающей их ткани обычно появляются аномальные медленные
волны и ЭЭГ можно использовать для уточнения локализации таких опухолей:- ЭЭГ позволяет судить, о функциональном состоянии коры, например, о глубине наркоза,
о наличии и определенных зонах патологических процессов.Было проведено бесчисленное количество исследований, направленных на поиски ЭЭГ-коррелятов интеллекта,
особенностей личности, поведения. Результаты в большинстве случаев оказались
разочаровывающими. В мозгу человека более 10 млрд нервных клеток, сплетенных в
плотную есть взаимными связями. Даже в самых тонких записях ЭЭГ неизбежно
выявляется лишь слитная трескотня сотен тысяч клеток, приглушенная и искаженная
черепом. По мнению одного из исследователей: "Мы подобны слепым, пытающимся
понять работу фабрики, прислушиваясь снаружи к се шуму" (Margcrison ct al., 1967).Изучение изменений ЭЭГ начинается с анализа частот и амплитуд мозговых волн. - альфа-ритм: при низкой активности мозга большие группы нервных клеток разряжаются одновременно. Эта синхронность
отображается на ЭЭГ в виде последовательности медленных волн с частотой от 8-13
гц, имеющих большую амплитуду. Альфа-ритм наиболее четко регистрируется главным
образом в затылочных отведениях, когда человек находится в расслабленном
состоянии, с закрытыми глазами. - бета-ритм: во время активной работы мозга каждая участвующая в ней нервная клетка разряжается в соответствии со своей
специфической функцией в своем собственном ритме. В результате активность
становится абсолютно асинхронной и регистрируется в виде быстрых волн высокой
частоты (выше 13 Гц) и малой амплитуды. Амплитуда бета-волн уменьшается по мере
увеличения активности мозга. Ранние наблюдения о связи этих волн с состоянием относительной активности были в общем подтверждены. - тета-ритм: колебания от 4 до 8 гц. Они появляются на первой стадии сна, а также у некоторых опытных мастеров медитации
или во время пребывания испытуемых в изолированной камере в условиях сенсорной
депривации.- дельта-ритм: колебания с частотой менее 4 гц. Регистрируется но время глубокого сна, а также при
некоторых патологических состояниях (опухоли мозга) или у больных незадолго до смерти.Следует подчеркнуть, что разбиение на группы по частоте более или менее произвольно - ононе соответствует каким-то физиологическим категориям. И связьсо степенью "психической активации" представляется довольно слабой,
так как есть много исключений. Напимер, у взрослых людей при эмоциональных
переживаниях обнаруживаются колебания гста-частоты. Кроме того, эта классификация не подходит для всех возрастов: альфа-ритм появляется только в раннем подростковом возрасте.При первых попытках более систематического анализа изменений ЭЭГ обычно измеряли и частоту, амплитуду.
Одна из наиболее часто используемых зависимых переменных - это "время
альфа-ритма", т.е. процент времени, занимаемый альфа-ритмом.Еще один традиционный показатель ЭЭГ - это "блокада альфа-ритма", т.е. внезапное очень резкое
уменьшение амплитуды альфа-волн, которое обычно происходит при предъявлении
раздражителей. Например, если на глаза испытуемого падает свет, то в затылочных
отделах мозга у него обычно происходит блокада альфа-ритма.Появление компьютерной техники способствовало появлению новых методов вычисления различных показателей ЭЭГ. В
частности, многие исследователи стали проводить частотно-амплитудный анализ
регистрируемых волн электрической активности мозга. По мере развития
компьютерных способов математической обработки ЭЭГ появлялись вес новые
возможности для изучения механизмов работы мозга.Так, электроэнцефалографические исследования Л.П. Павловой (1988) обнаружили для
разных видов деятельности человека универсальное правило "смещения фокуса
максимальной активации (ФМА). Например, при выполнении сознательных
целенаправленных действий ФМА устойчиво регистрируется в речевых зонах левого
полушария. Затем возникает генерализация возбуждения и распространение ФМА на
ряд других корковых зон. Другими словами, наблюдается смена
доминантно-субдоминантных отношений между полушариями и лобными и теменными
ассоциативными областями коры. Завершающаяся перестройка активности коры
сводится к устойчивому смещению ФМА в правое полушарие, а затем в задние отделы
коры. В лобных долях на этой стадии ФМА наблюдается в течение коротких
промежутков времени. Наиболее выраженное смещение ФМА обнаружено в процессе
предметной (внешне развернутой) деятельности и в процессе мыслительной
(свернутой) деятельности. Интеллектуальные психические процессы всегда вовлекают
в активацию лобные доли, но не обязательно левого полушария. При автоматизации
интеллектуального навыка наблюдается значительное усиление альфа-ритма в левой речевой зоне Брока, а ФМА смещается в правые лобные области. Такая
активность мозга соответствует оперативному наглядно-действенному мышлению.
Динамика перестройки корковой активности оказалась принципиально схожей при
изучении разных видов деятельности: перцептивной (задачи на опознание),
познавательной, (мышление), коммуникативной (речевое общение),
преобразовательной (предметно-развернутые виды деятельности).Кроме того, обнаруживаются индивидуально-устойчивые, привычные (в покос и при работе) типы корковой
активации. А.А. Ухтомский подчеркивал важность изучения
индивидуально-личностных особенностей "мозгового хронотопа", свойств
интроверсии (аутизма), способностей к дифференциальной срочности реакций,
разных способов оценки ситуаций: либо по пути длительного логического
сукцессивного анализа, либо путем "внезапной рецепции зорким глазом
специалиста" сразу всей обстановки, то сеть симультанном охватывании
существенных сторон ситуации. Он говорил об индивидуально выраженных
способностях к различению существенного от несущественного в потоке ближайших
конкретных ощущений с адекватными рефлексами на них, указывал изучать тс пути и
средства, которые приводят к развитию таких способностей в масштабах истории.Была заложена, таким образом,основа для индивидуального различия когнитивных (познавательных) стилей.
Совокупность индивидуальных особенностей психики и поведения человека
составляет тип высшей нервной деятельности, или темперамент человека. Он
складывается из общих свойств нервной системы, которые характеризуются: 1)
экстра-интроверсией; 2) -эмоциональной стабильностью-невротизмом и 3)
подвижностью или инертностью нервных процессов. Проблема психофизиологических
основ индивидуально-типологических различий людей становится наиболее
актуальной научно-практической задачей.
Почему вы так волнуетесь? У вас прекрасная болезнь, болезнь Пушкина.
|
Статус: нет меня
|
|
|
|
John_Smith
|
#28 | Среда, 02.07.2014, 12:12
|
Автор темы
Фильм
Юзер-бар +
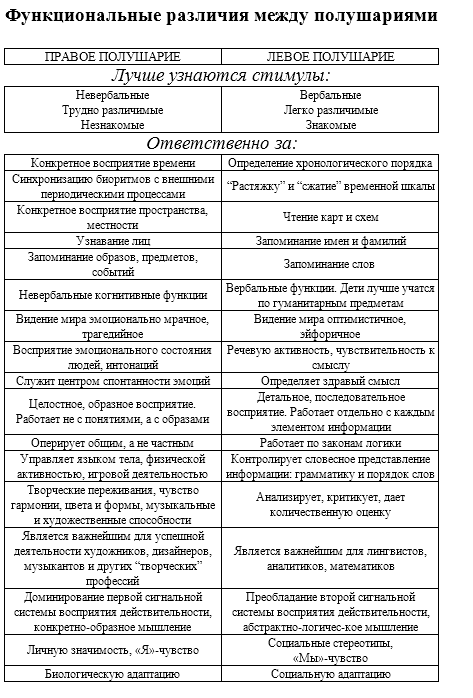
Почему вы так волнуетесь? У вас прекрасная болезнь, болезнь Пушкина.
|
Статус: нет меня
|
|
|
|
John_Smith
|
#29 | Среда, 02.07.2014, 12:13
|
Автор темы
Фильм
Юзер-бар +
Локализация функций в коре большого мозга
Мозг - это ведущий орган организма, каждый человек - это его мозг. Многие из наших современных представлений о функциях мозга восходят к Галлю - отцу френологии. В начале XIX века Галль провозгласил,
что мозг человека состоит из 27 отдельных органов, каждый из которых ответственен за одну из наших основных способностей. Например,можно ожидать, что обладатель шишек на левом виске будет склонен к стяжательству. Идеи Галля в дальнейшем неподтвердились. Однако он зародил зерно мысли о локализации функций в мозгу.
Хотя при нашем уровне знаний уже невозможно представить себе, что каждая отдельная
область мозга управляет отдельным видом поведения, мы, бесспорно, можем
говорить по крайней мере об известной степени локализации мозговых функций.
Например, анатомические связи, идущие от глаз, образуют четко обособленный путь
от сетчатки к области в затылочной коре. Одна из главных задач современных
исследований мозга - попытаться выяснить, в какой степени локализованы функции
различных типов.За полтора столетия, прошедшиепосле работ Галля, представление о локализации функций несколько раз входило в
фавор и впадало в немилость. Исследования Пьера Флуранса в начале 19 века говорили
в пользу холистического взгляда. На основе наблюдений, которые оказались
ошибочными, он сделал вывод о диффузном представительстве сложных психических
функций в мозгу. Эта концепция "эквипотенциальности" различных
участков мозга властвовала в западной медицине вплоть до 1861 года, когда Поль
Брока на заседании Парижского антропологического общества представил результаты
своих наблюдений над больными с повреждениями мозга. Он продемонстрировал
препараты мозга нескольких больных, которые после апоплексии потеряли
способность говорить и у которых были ограниченные повреждения в левом полушарии.Сходные клинические данныеВернике и других исследователей воскресили идею о локализации функций в мозгу.
Примерно в тот же период теория локализации получила подкрепление в
экспериментах Фрича и Гитцига, показавших, что электрическое раздражение
определенных участков коры вызывало у собаки совершенно определенные мышечные
движения.В 1905 г. Пьер Мари повторно исследовалпрепараты, представленные Брока, и показал, что его выводы тоже были основаны
на ошибочных наблюдениях: поражения мозга были значительно более обширными, чем
думал Брока. В итоге снова поддержку получила холистическая точка зрения.Обширные исследования КарлаЛешли, который в первой половине этого столетия изучал влияние повреждений
мозга на результаты научения крыс в лабиринте, также подкрепляли холистическую
точку зрения. Данные Лешли указывали на то, что ухудшение функций зависит от
общего объема разрушенной мозговой ткани, а не от повреждения специфических
участков. Лешли абсолютизировал представление об эквипотенциальности - любая
часть коры, утверждал он, может принять на себя функцию любой другой. Однако
даже он постепенно пришел к признанию того, что в коре есть некоторое
"разделение труда".Сегоднявопрос состоит не в том, существует ли локализация функций в мозгу человека, а
в том, насколько сильно она выражена. Один из наиболее авторитетных современных
исследователей в этой области - советский нейрофизиолог А.Р. Лурия (1973) -
выдвинул представление о том, что мозг организован как система функциональных
единиц, координирующих сложные психические процессы. Каждая такая единица
связана с определенными участками коры. Другие исследователи продолжают придерживаться
мнения о большей или меньшей степени локализации.Именнов этой области нейробиологии уникальный вклад могут внести исследования ЭЭГ,
которая отражает функционирование мозга здорового человека. Как мы уже указывали,анатомически кору обычно разделяют на четыре главных отдела, или доли.
Функционально кору разделят на зоны. В коре различают: сенсорные
(чувствительные), моторные (двигательные) и ассоциативные зоны.Сенсорные зоны. Разные сенсорные модальности представлены в долях полушарий по-разному.
Соматосенсорная зона – это область проприоцептивной, кожной и висцеральной чувствительности, располагающаяся в задней центральной извилине. При ее
раздражении возникает ощущение прикосновения, покалывания или онемения. Иногда
возникает ощущение низкой или высокой температуры, иногда - ощущение боли.
Сенсорная зрительная зона располагается в затылочной области коры. В этузону приходят нервные импульсы от рецепторов сетчатки, здесь происходит обработка
зрительных стимулов. При раздражении этой зоны возникают простейшие зрительные
ощущения: вспышки света, наступления темноты, различные цветовые ощущения. При
этом никогда не возникают сложные зрительные образы.
Сенсорная слуховая зона располагается в височном отделе коры. К ней приходятчувствительные пути от рецепторов слухового аппарата. Раздражение этой области
вызывает ощущение низких или высоких, громких или тихих звуков. При этом
никогда не возникает ощущение мелодии или речевых звуков.
Зона вкусовых ощущений располагается в теменной области, точнее в нижней части задней центральной извилины. К ней приходят импульсы от вкусовых
рецепторов полости рта и языка. При раздражении этой зоны возникают различные
вкусовые ощущения.
Моторные зоны коры – участки коры больших полушарий при раздражении которых наблюдаются различные движения. Они взаимодействуют с
сенсорными зонами, вследствие чего при раздражении некоторых участков моторных
зон вместе с движениями возникают и ощущения. Моторные зоны расположены впередней центральной извилине. При ее поражении наблюдаются серьезные нарушения
движений у человека.
Ассоциативныезоны коры – участки коры, расположенные рядом ссенсорными зонами. Причем при поступлении импульсов в сенсорные зоны возбуждение
возникает и в ассоциативных зонах. Другой их особенностью является то, что в
ассоциативной зоне возбуждение может возникать при поступлении импульсов от
различных рецепторов. Здесь образуются ассоциативные связи между специализированными
сенсорными областями и интегрируется приходящая из них информация. Кроме того,
здесь, как полагают, текущая информация объединяется с эмоциями и
воспоминаниями, что позволяет людям думать, решать проблемы, составлять планы.
Неслучайно, что эти зоны наиболее хорошо развиты у человека. Показано, что ассоциативные полятеменной доли объединяют информацию, приходящую от соматосенсорной коры
(сообщения от рецепторов кожи, мышц, сухожилий и суставов) относительно
положения тела и его движений, со зрительной и слуховой информацией,
поступающей из зрительной и слуховой коры. Эта объединенная информация
позволяет нам иметь точное представление о собственном теле во время
передвижений в окружающем пространстве.Информация, интегрированная в теменнойассоциативной зоне, передается затем в лобную кору. Благодаря обширным
двусторонним связям с лимбической системой к этой картине добавляются эмоции, а
также информация, взятая из памяти. Другие нервные связи доставляют в лобный
ассоциативный отдел информацию, позволяющую оценить текущие требования
организма и окружающей среды и выбрать среди них первоочередные – решить, чтолучше, а что хуже для организма в данной ситуации. Лобная кора, по-видимому,
ответственна и за выбор целей, которые человек ставит перед собой, а также за
нашу оценку различных обстоятельств связи с этими целями.Сведения о людях с повреждениямилобных долей действительно подтверждают решающую роль этих областей коры в
выработке суждений и построении планов. Лица с такими повреждениями испытывали
огромные трудности в приспособлении к жизненным конфликтам и меняющимся
требованиям. Они становились безответственны, не способными вести себя так, как
этого требовала ситуация, или осуществить сколько-нибудь последовательные
жизненные планы.Представления о зонах коры былисформулированы на основе идей И.П. Павлова, который рассматривал кору большого мозга как совокупность корковых
концов анализаторов. По его мнению, в корковом конце различают “ядро” сенсорной
системы и “рассеянные элементы”. Ядро – это участок коры, в который точно
проецируются все периферические рецепторы. Рассеянные элементы располагаются
вблизи ядра и на различном расстоянии от него. В ядре, предположительно,
оуществляется более простой анализ поступающей информации, а в рассеянных
элементах – более сложный анализ и синтез информации. При этом зоны рассеянных
элементов различных анализаторов не имеют четких границ и могут наслаиваться
друг на друга. Корковые концы анализаторов,осуществляющие анализ и синтез сигналов, поступающих из внешней и внутренней
среды организма, составляют первую сигнальную систему действительности.В отличие от первой, втораясигнальная система имеется только у человека и она тесно связана с развитием
речи. Считается, что речь и мышление осуществляется при участии всей коры. В то
же время, имеются зоны коры, являющиеся центрами целого ряда специфических
функций, связанных с речью. Так, двигательные анализаторы устной и письменной
речи располагаются в области лобной доли коры, прилежащей к передней
центральной извилине. Анализаторы зрительного и слухового восприятия речи
находятся вблизи ядер анализаторов зрения и слуха.
Причем у большинства людей
(праворуких и леворуких) центры речи локализованы в левом полушарии.Обозначим локализацию основныхядер анализаторных систем в коре полушарий:^
Ядродвигательного анализатора – передняя центральная извилина. Здесь происходит
анализ сознательных проприоцептивных (мышечно-суставных) ощущений и формируется
кортикоспинальный тракт.¨
Ядроанализатора кожной чувствительности – задняя центральная извилина.¨
Ядроцентра стереогнозии (узнавание предметов на ощупь) – верхняя теменная долька.¨
Ядроцентра апраксии (praxis– практика) – способности производить целенаправленные координированные
движения – надкраевая извилина.¨
Ядродвигательного центра письменной речи (центра графии) – задний отдел средней
лобной извилины.¨
Ядроцентра артикуляции речи, центр Брока (способность членораздельно произносить
слова) – задний отдел нижней лобной извилины.¨
Ядрозрительного анализатора – находится в верхней части затылочной доли.¨
Ядрокорково-зрительного анализатора, письменной речи – угловой извилиной.¨
Ядрослухового анализатора – задний отдел верхней височной извилины.
Ядрокорково-слухового анализатора речи – средняя треть верхней височной извилины
Почему вы так волнуетесь? У вас прекрасная болезнь, болезнь Пушкина.
|
Статус: нет меня
|
|
|





